高原湿地的生命支柱和主宰
高原湿地土壤类型 高原湿地土壤主要为沼泽土,属于隐域性土壤类型,是在寒冷湿润的环境和沼泽化草甸植被条件下发育而…
高原湿地土壤类型 高原湿地土壤主要为沼泽土,属于隐域性土壤类型,是在寒冷湿润的环境和沼泽化草甸植被条件下发育而…
湿地的存在以水为特征,水是湿地生生不息的脉搏。青藏高原的高山冰雪融水是河流的主要水源,是东亚和南亚地区诸多江河…
高寒气候是高原湿地形成的驱动力,也是高原湿地形成的基础。因为水分与热量直接影响着沼泽植物种类组合及其生长发育、植物残体的分解量、分解强度和泥炭物质聚集。高原湿地生态系统在漫长的气候条件变动的情况下,不断发生迁移与演化。新生代以来,全球和青藏高原地区发生的多次冷暖交替,冰期和间冰期的反复出现,以及高原本身多次抬升与夷平的相互交替,成为湿地生态系统演化的重要驱动力。
青藏高原具有气候地带性特征。气温和降水随海拔高度的增加而变化,在水平方向上自南向北可以划分出热带、亚热带、高原温带、高原亚寒带和高原寒带等气候带。青藏高原的动力和热力作用是造成高原独特气候的重要原因,其中太阳辐射是青藏高原动力和热力作用的主要能量来源。它不仅对该地区气温、气候和天气类型有重要影响,而且是形成高原特有生态现象的主要能量来源。青藏高原太阳总辐射量是全国太阳总辐射值最高的地区。太阳总辐射量与年降水有一定的关系。一般降水季节,云量多,太阳总辐射偏少,气温日较差大,年较差小。最高气温分布在雅鲁藏布江大拐弯以南地区和横断山区的三江流域地区,年均气温分别为 18℃和 12℃;气温最低的地方分布在高原内部,藏北高原、巴颜喀拉山的玛多和清水河以及祁连山的托勒,1 月平均气温低于-18℃,极端最低温度为-41℃。青藏高原上降水的分布受西南季风的影响, 不同地区降水量相差悬殊,最大降水量地区的降水量与最小降水量地区的降水量相差 200 ~ 300 倍。西藏东南部的平均降水量在 4000mm 以上,其中巴昔卡年降水量达 4500mm,是我国降水最多的中心之一。由此逐渐向高原西北部地区减少,到柴达木盆地的西北部年降水量只有 200mm 左右。整个藏北高原年平均降水量仅为 300 ~ 500mm。由此可见,高原各地区降水季节分配极不均匀,雨季和旱季分异非常明显。除喜马拉雅山南坡雨季(6 ~ 9 月)降水量占年降水量的 60%左右外,高原其他地区一般雨季降水量占年降水量的 90%左右。高原降水的日变化也非常明显,降水常常伴随着夜幕降临悄然而至,夜雨率达 50%以上。科学研究表明夜雨有利于各种植物的生长。
青藏高原寒冷湿润的气候特征造就了大面积的高寒湿地,其中若尔盖高原以及长江、黄河、怒江等江河源区的湿地分布面积最大。
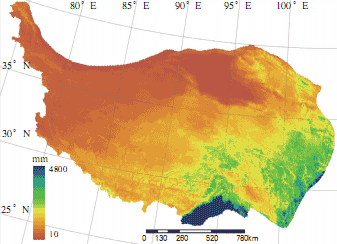 青藏高原多年平均降水分布图
青藏高原多年平均降水分布图
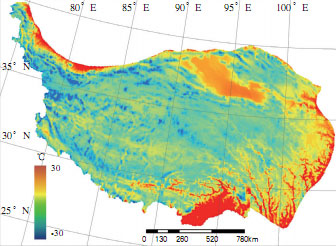
青藏高原多年平均气温分布图
山地地貌为高原湿地的形成和发育提供了生存空间,一般低洼地区(如盆地、洼地)比丘陵和高地更有利于沼泽发育,因为负地貌有利于水的汇聚,不利于水的排泄,成为湿地发育的理想场所。不同地貌类型对湿地的形成和发育有着不同的影响。
山地、高原是我国沼泽发育的重要地貌类型。我国是一个多山的国家,山地、丘陵约占国土面积的 2/3。全国地势西高东低,形成三大阶梯。 西南部耸立着号称世界屋脊的青藏高原, 平均海拔在4500m以上,为我国第一级阶梯;中部为由内蒙古高原、黄土高原、云贵高原和一系列高大山地组成的第二级阶梯;在大兴安岭、太行山、巫山、雪峰山以东,主要是由东北平原、华北平原、长江中下游平原和低山丘陵为主构成的第三级阶梯。
位于第一级阶梯的青藏高原地貌类型多样, 既有高大的山脉, 又有高原湖盆、 山原湖盆、谷地和高山深谷等多种特殊的地貌类型。青藏高原面及其四周边缘有一系列东西向和南北向巨大高山山脉。东西向山脉占据了青藏高原的大部分地区,从北到南有阿尔金山-祁连山、昆仑山、巴颜喀拉山、喀拉昆仑山、唐古拉山、冈底斯山、念青唐古拉山和喜马拉雅山。除祁连山山顶海拔在 4500 ~ 5500m 以外,其余山脉山顶都在海拔6000m以上。南北向山脉主要分布在高原东南部和横断山地区,自西向东有伯舒拉岭、他念他翁山、宁静山、大雪山和龙门山-夹金山-大凉山。这些山脉山顶海拔多在4500~7000m。这些山脉成为整个青藏高原地区的地貌骨架,控制着高原地区地貌的基本格局,进而也控制着整个高原地区生态系统的空间分布格局。青藏高原面为小起伏高山、高海拔丘陵和宽谷盆地的组合体,小起伏高山和高海拔丘陵为不同时代的地形面,而宽谷盆地主要为第四纪的堆积面。在青藏高原形成过程中,因后期内外营力的作用,使高原面有不同程度的变形,整个高原地势由西北向东南倾斜,地处腹地的羌塘高原面保存较好,而处在高原边缘的横断山脉地区为残留的平坦山顶面。山地剥蚀形成的夷平面地表平坦,排水不畅,深厚的风化壳阻碍水分下渗,造成地表过湿或发生积水,形成有利于沼泽发育的环境。横断山区一些沼泽发育在第三纪形成的夷平面上。山地、高原中多发育封闭或半封闭的山间盆地,盆地内地势平坦,排水不畅,且多有地下水出露补给盆地,使盆地内汇集地表水和地下水,发生沼泽化过程。如青藏高原东北隅的若尔盖高原就是川西北高原内发育的断陷盆地。由于高原盆地内湿地形成发育的环境比较稳定,湿地发育条件能够长期保持,因而发育泥炭沼泽并积累较厚的泥炭层。
冰川地貌和冻土地貌是沼泽形成、发育的有利地貌。青藏高原、长江、黄河河源区沼泽的形成、发育多与冰川、冻土地貌有密切关系。冰川,尤其是大陆冰川退缩后,在原冰川发育区留下了一系列冰蚀、冰积地貌,具体地貌类型为冰斗、围谷、槽谷、冰蚀洼地等低洼地貌。这些地貌底部堆积大量冰碛物,透水性能不好,形成隔水层,成为湿地形成发育的温床。青藏高原上现代冰川分布广泛,冰川地貌普遍发育,现有冰川面积达 1168.18km2,特别是唐古拉山北坡有各类山谷冰川 520 条,占青藏高原冰川总数的 82.9%,冰川一方面塑造有利于沼泽发育的地貌,另一方面又以融水补给冰川洼地使地表长期稳定积水或土壤过度湿润,从而发育沼泽。冻土层形成的区域性隔水层阻碍了冰雪融水的下渗,冻融作用又使地表形成冻融洼地,有些地方冻土融化后,地表下陷形成热融湖,在这些谷地洼地、热融湖区经常有积水或土壤过湿,以致发生区域性沼泽化现象。在冻土地貌区沼泽分布十分广泛,不仅在河漫滩、阶地上发育沼泽,在整个宽阔的浅平谷地中,甚至分水岭上都有沼泽。
地质构造是地貌发育的基础,地壳运动造成的大规模褶皱、断裂等主要构造形态以及与之相伴的隆起和坳陷,控制着地貌的主要发展方向和平面格局,进而影响湿地的形成和发育。大断裂带之间或两组断裂带之间往往有利于湿地的形成和发育。因为断裂带之间容易发育成断陷盆地,在盆地内常常形成较平坦的沉积平原,有利于地表水汇集,形成湿地。地壳运动的形式对湿地的形成发挥着至关重要的作用。新构造运动缓慢下沉时,或保持相对稳定的速度下沉时, 对地面的侵蚀作用减弱, 在地面平坦或低洼地段, 由于不利于把水分排出去,形成地表过湿或薄层积水的环境,促使湿地成长发育;而当新构造运动上升时,由于强烈的侵蚀作用,导致地下水位下降,地表自然疏干,不利于湿地的形成。
青藏高原平均海拔 4000m 以上,是我国大陆地势上最高的一级台阶,也是地球上最高的一级地貌台阶,有“世界屋脊”之称。它既是一系列巨大山系和辽阔的高原面的组合体,也是近 300 万年来大面积强烈隆起的巨大的构造地貌单元。4000 万年以前,在现今青藏高原范围的南部还是一片汪洋大海,科学命名为古地中海,又名特提斯海(洋) 。目前对当时这一海洋的大小还有不同的认识。近年来根据新的研究成果,这个洋的深水部分宽度至少在 1500km 以上,如果再加上两侧宽阔的大陆架,洋面总的宽度可达数千千米。在北部,那时还有一些残留的海湾伸向陆地内部,如昆仑山北缘、塔里木盆地的西南缘和南天山的西南缘。可见高原北部当时虽已是陆地,但海拔并不高,濒临海洋,河流和湖泊纵横交错,属濒海平原到丘陵盆地面貌。那时青藏高原还没有形成,西藏南部还被海水所淹没着。根据古地磁的研究成果,现在的喜马拉雅地区当时还处于赤道附近,在 4000 万年前快速向北漂移,才与亚洲大陆碰撞相连接,青藏高原这块陆地从此诞生了。地球已有 46 亿年的历史了,而青藏高原地区仅在 4000 万年以来才形成统一的陆地,后经逐步抬升才形成我们今天所看到的高原面貌。由此可见,青藏高原形成于地球历史最晚时期的今天,她的成长历史还十分短暂而年青,直到现在还在继续发育成长。
青藏高原在近几百万年以来经历了大幅度强烈隆起抬升,其间也有相对沉降区,形成许多较大的山原宽谷地,如高原东南部的长江、黄河河源区,四川西北高原中的若尔盖山原宽谷区,以及那曲山原宽谷区等,这些新构造运动下降区是青藏高原湿地的重要分布区。新构造运动对青藏高原湿地的控制作用以若尔盖高原湿地最为典型。
在区域大规模上升运动中,沿老断裂带发生区域性断陷,而后又发生不均衡的相对下降而形成的昆明构造盆地以及黔西一些构造——岩溶盆地,这些地区都是沼泽的广泛发育区。
鸟类物种的中文名称中含有一些生僻字,往往被错读或不见于新华字典。东北林业大学常家传教授对鸟名中的生僻字总结如下…
鸟类迁徙是自然界中最引人注意的生物学现象之一,世界上每年有几十亿只候鸟在秋季离开它们的繁殖地,迁往对它们更为适宜的栖息地。迁徙路线就是候鸟在迁徙过程中使用的一条宽大通道,现已知全世界有 8 条主要候鸟迁徙路线。在欧洲和亚洲共有 5 条主要迁徙路线,即东亚—澳大利西亚、中亚—印度、西亚—非洲、地中海—黑海、东大西洋迁徙路线。中国湿地水鸟的迁徙路线主要分为东部、中部和西部 3 条候鸟迁徙路线,其中东部迁徙路线是东亚—澳大利西亚迁徙路线的重要组成部分,西部迁徙路线为中亚—印度迁徙路线的重要组成部分。
(1)东部迁徙路线是我国湿地水鸟最重要的迁徙路线。在俄罗斯、日本、朝鲜半岛和我国东北与华北东部繁殖的湿地水鸟,春、秋季节主要通过我国东部沿海地区进行南北方向的迁徙。春季,来自南洋群岛和大洋洲的北迁鸟类到达台湾后,分为两支,一支沿中国大陆扩散或继续沿东部海岸北上,另一支经琉球群岛到日本或继续北迁。沿中国大陆东部沿海北迁的鸻鹬类等湿地水鸟在到达长江口以后, 又分两条北上迁徙路线。一条经江苏、 山东到东北、俄罗斯,另一条则越海向朝鲜半岛或日本迁飞。秋季,湿地水鸟沿中国东部沿海向南迁飞至华东和华南,远至东南亚各国,或由俄罗斯东部途经中国向东南亚至澳大利亚迁徙,其南下迁徙路线大致与春季北上路线相似。
(2)西部迁徙路线。内蒙古西部、甘肃、青海和宁夏的湖泊、草甸等湿地繁殖的候鸟,在秋季可沿阿尼玛卿山、巴颜喀拉山和邛崃山脉向南迁飞,然后沿横断山脉南下至四川盆地西部和云贵高原越冬,有些候鸟可飞至中南半岛越冬。新疆地区的湿地水鸟可向东南汇入该西部迁徙路线,或向西南出境,或向南进入西藏。西藏地区的湿地水鸟主要沿唐古拉山和喜马拉雅山向东南方向迁徙,亦可以飞越喜马拉雅山脉,至印度、尼泊尔等地越冬。
(3)中部迁徙路线是我国境内的一条迁徙路线。在内蒙古东部、中部草原,华北西部和陕西地区繁殖的候鸟,秋季沿黄河流域、吕梁山和太行山南下,越过秦岭和大巴山区进入四川盆地越冬;或继续沿大巴山东部向华中或更南的地区越冬。湿地水鸟在迁徙的过程中,一般只有少数种类可以不间断地飞行完成其整个迁徙过程,大多数鸟类会在沿途的某些被称之为停歇地的地方停下休息和补充能量。因此,迁徙停歇地对湿地水鸟完成其迁徙过程起着极为重要的作用。
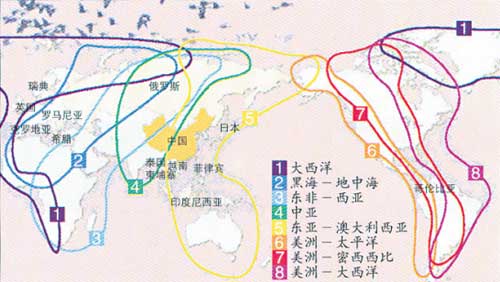
全球候鸟迁徙路线(引自湿地国际,2006)
根据湿地水鸟地理分布特征,根据中国自然地理环境的地域差异和湿地类型的不同,我国湿地水鸟栖息地可分 5 种类型,即东北沼泽湿地大型涉禽和游禽繁殖与迁徙停歇区、西北和青藏高原草甸沼泽与高原湖泊游禽和大型涉禽繁殖与迁徙停歇区、西南部高原湖泊和湿草甸水鸟游禽和大型涉禽越冬与迁徙停歇区、长江中下游淡水湖泊大型涉禽和游禽越冬区、沿海和近海岛屿滨海滩涂湿地 鹬目鸟类与大型涉禽繁殖越冬与迁徙停歇区。
(1)我国东北地区是国内最大的淡水沼泽湿地区,有 6 处被列为国际重要湿地,分别是内蒙古达赉湖,黑龙江洪河、兴凯湖、三江和扎龙,吉林向海等自然保护区。这一地区的湿地水鸟种类丰富,且特别适合丹顶鹤、白头鹤、白鹤、白枕鹤、灰鹤、白鹳、东方白鹳、黑鹳、黑头白鹮、白琵鹭等大型涉禽和大天鹅Cygnus cygnus、中华秋沙鸭、遗鸥等游禽的栖息繁殖,是亚洲东北部湿地水鸟繁殖中心和亚洲北部湿地水鸟南迁的必经之地。例如著名的扎龙自然保护区内,就分布有丹顶鹤、白枕鹤、白头鹤、蓑羽鹤、白鹤、灰鹤等 6 种鹤类,且为丹顶鹤的最重要繁殖地。
(2)我国西北部与青藏高原地区分布众多高原湖泊、沼泽、草甸等类型的湿地。有内蒙古的鄂尔多斯遗鸥自然保护区,青海的青海湖鸟岛自然保护区、扎陵湖湿地、鄂陵湖湿地,西藏麦地卡湿地、玛旁雍错湿地等 6 处被列为国际重要湿地。每年有黑颈鹤、大天鹅、斑头雁Anser indicus、赤麻鸭Tadorna fer-ruginea、黑鹳、遗鸥、渔鸥Larus ichthyaetus、棕头鸥L. brunnicephalus、红嘴鸥L.ridibundus、红脚鹬Tringa totanus、普通鸬鹚等大量湿地水鸟在该地区栖息繁殖。例如,鄂尔多斯遗鸥自然保护区是遗鸥最重要的繁殖地,其种群数量占世界遗鸥繁殖种群的 60%;新疆的巴音布鲁克自然保护区为大天鹅的重要繁殖地;地处西藏南羌塘高寒湿地的雅鲁藏布江中游河谷黑颈鹤自然保护区是黑颈鹤最重要的繁殖地,其数量可占黑颈鹤繁殖种群的80%;青海湖鸟岛自然保护区是斑头雁、棕头鸥、渔鸥和普通鸬鹚的重要繁殖地;另外,塔里木河流域是我国黑鹳的重要繁殖地。
(3)我国西南部的云贵高原既有高原湖泊,又有草甸,是内陆迁徙湿地水鸟的重要越冬地和迁徙停歇地。如云南大山包黑颈鹤自然保护区、纳帕海自然保护区、拉市海自然保护区、碧塔海自然保护区等 4 处为国际重要湿地。云贵高原是黑颈鹤的最重要越冬地,其中大山包黑颈鹤自然保护区是最大越冬地;贵州草海自然保护区不仅是黑颈鹤的重要越冬地,而且还是灰鹤、白头鹤、黑鹳、凤头、白琵鹭、灰雁Anser anser、斑头雁、赤麻鸭等鸟类的重要越冬地;在拉市海湿地每年有 10 多万只候鸟栖息与停歇,越冬湿地水鸟数量达 30 000只 ;而在纳帕海湿地仅雁鸭类就有近万只。
(4) 长江中下游地区的湿地以大型的淡水湖泊为主,分布有许多著名淡水湖泊,如洞庭湖、鄱阳湖、升金湖、太湖、龙感湖、洪泽湖、高邮湖、巢湖和沉湖等。湖南的东洞庭湖自然保护区、南洞庭湖湿地和水禽自然保护区、汉寿西洞庭湖自然保护区和江西的鄱阳湖自然保护区等 4 处被列为国际重要湿地。该地区不仅湖泊众多,而且每年秋冬季节湖泊水位下降还可形成大面积的浅滩沼泽,从而成为白鹤、白枕鹤、白头鹤、东方白鹳、黑鹳、中华秋沙鸭和各种雁鸭类等数量庞大的湿地水鸟主要集中越冬地和繁殖地。例如,中国最大的淡水湖泊鄱阳湖不仅是白鹤、白头鹤和灰鹤的重要越冬地,而且还有大群的大天鹅、小天鹅Cygnus columbianus、鸿雁、豆雁A. fabalis、白额雁A. albifrons、小白额雁、灰雁、赤颈鸭A. pe-nelope、罗纹鸭A. falcata、赤膀鸭A. strepera、绿翅鸭A. crecca、绿头鸭A. platyrhynchos、斑嘴鸭A. poecilorhyncha、针尾鸭A. acuta 等雁鸭类,以及卷羽鹈鹕、黑鹳和东方白鹳等湿地水鸟在此越冬。
我国沿海和近海岛屿地区有大量的滨海滩涂湿地。辽宁双台河口自然保护区、江苏盐城沿海滩涂珍禽自然保护区、大丰麋鹿自然保护区、上海市崇明东滩鸟类自然保护区、广西山口红树林自然保护区、广东湛江红树林自然保护区、海南东寨港自然保护区和香港米埔—后海湾等 8 处被列为国际重要湿地。我国沿海和近海岛屿地区既是大量湿地水鸟的极重要迁徙中途停歇地,又是许多湿地水鸟的重要越冬地和繁殖地。例如,双台子河口湿地是黑嘴鸥的重要繁殖地;山东半岛的荣城海滨为我国北方沿海大天鹅的主要越冬地;盐城沿海滩涂湿地几乎集中了中国境内的所有丹顶鹤越冬群体,并有大量鹭类和雁鸭类在此越冬;上海崇明东滩湿地为鸻鹬类极重要的迁徙停歇地;浙江温州沿海滩涂是黑嘴鸥的最重要越冬地;台湾曾文溪口滩涂是世界最大的黑脸琵鹭越冬种群栖息地;香港米埔—后海湾湿地是我国南部十分重要的湿地水鸟越冬地和迁徙中途停歇地,每年有数以万计的鸻鹬类、鸥类、鹭类、鸭类以及黑脸琵鹭、黑嘴鸥、勺嘴鹬、小青脚鹬、半蹼鹬Limnodromus semipalmatus 等鸟类在此地越冬。
中国分布有湿地水鸟 262 种,其中潜鸟目4 种,目5种,鹈形目8种,鹳形目 34 种,红鹳目 1 种,雁…
由于湿地类型的不同和湿地水鸟适应方式的差异,湿地水鸟在长期的进化过程中逐渐形成了不同的生态类型,其中主要有游禽和涉禽等两种类型。
(1)游禽 是适应在水中游泳、 潜水捕食生活的鸟类。 包括潜鸟目、目、鹱形目、 鹈形目和雁形目的所有种类, 以及鸥类。其中鹱形目和鹈形目的鹲科、鲣鸟科和军舰鸟科为远洋性海鸟;雁形目的鸭、雁和天鹅等往往被人们称之为水禽。由于鸥类在生活习性上适应游泳,有学者认为应归入游禽类;但又由于鸥类在形态和演化关系上与鸻鹬类相近, 也有学者将其归为涉禽。可见,鸥类是介于游禽和涉禽之间类型的湿地水鸟,那就称其为善游泳涉禽和善涉行游禽。
游禽能在各种类型的水域活动,从海洋到内陆河流、湖泊都有游禽的身影,有的在浅水滩觅食,有的在一定深度的水域潜水觅食。游禽多喜群居,经常成群活动。体型大小相差悬殊,如卷羽鹈鹕Pelecanus crispus 长可达180cm,而小Tachybaptus ru?collis 体长只有20cm左右。
游泳和潜水是游禽在水中的主要活动形式,它们的羽毛往往厚而致密、绒羽发达,构成有效的保暖层;多数种类的尾脂腺能分泌大量油脂,用喙涂抹在羽毛表面可用来防水。游禽的腿短而侧扁并从身体中央后移,后移程度可反映其潜水能力。一般腿越偏向身体后部的潜水能力越强,潜水越深;反之则不善潜水。游禽趾间具蹼,在游泳或潜水时,双脚直伸至尾后划动,有如船桨。游禽蹼的发达程度存在种类差异,如普通鸬鹚Phalacrocorax carbo四趾间均有蹼相连,叫全蹼足;鸭雁类是三趾间有蹼,称为常态蹼足;鸥类的趾间蹼不是很发达,称为凹蹼足;而像 的各趾两侧均有莲花瓣状的皮褶,称为瓣蹼足。
游禽的嘴型与其食性或捕食方式相关,有的嘴尖而具利钩(如鹈鹕和鸬鹚) ,有的在嘴缘有成排的锯齿(如秋沙鸭) ,使其捕鱼时防止猎物滑落;有些种类(如雁鸭类)具扁嘴, 且在嘴喙有成排的栉板, 用以滤食小型水生生物。游禽一般在近水区域营巢,巢比较简陋,有的在水面上营浮巢(如类) ,有的在水边地面以绒羽和草茎筑巢(如雁鸭类) ,有的在树上、灌丛和岩缝中用树枝营编织巢,并垫以羽毛和草茎等(如鹭类、鸬鹚和鹈鹕) ,有的直接产卵在岩石洞穴内(如鹱形目鸟类)或甚至直接产卵在岩石和地面上(如潜鸟类和燕鸥类) 。许多游禽会选择在岛屿筑地面巢,多成群且地点固定,往往形成非常壮观的鸟岛。鹱形目、鹈形目、鹳形目和鹤形目的绝大多数鸟类的雏鸟属于晚成鸟;而潜鸟目、目、雁形目和鸥类的雏鸟属于早成鸟。
(早成鸟:雏鸟孵出时发育较充分,被有密绒羽,眼已张开,腿脚有力,在绒羽干后,即可随亲鸟觅食。)
(晚成鸟:雏鸟出壳时尚未发育充分,体表光滑或微具稀疏绒羽,眼不能张开,不能独立生活,由亲鸟喂食,待在巢内完成发育后才能独立生活。)
游禽大多有迁徙的行为,且多数在北半球繁殖,并于每年的秋季集结成大群南迁到比较温暖的水域越冬,至翌年春季再返回北方的繁殖地。其中雁形目鸟类常常做南北向跨越大陆的迁徙,鹱形目鸟类沿赤道地区做东西向迁徙,而鸥形目则往往沿大陆海岸做跨越海洋的迁徙。
(2)涉禽?是一类适应于在浅水或岸边栖息生活的鸟类。包括鹳形目,红鹳目,鹤形目的鹤、秧鸡、秧鹤、日与日鳽 等科和鸻形目(鸥类除外)的所有种类,均为湿地水鸟。涉禽体型大小相差悬殊,大者如赤颈鹤Grus anti-gone, 体长可达170cm以上, 小者如小滨鹬Calidris minuta, 体长只有14cm左右。涉禽虽然在水边生活,但游泳能力一般,善于飞行,且姿态优美。为了适应涉水捕食的生活习性,涉禽的嘴、脚和颈部比其他生态类群的鸟类显著更长。腿长适于涉水,腿的长度与其能够涉水的深度直接相关。涉禽的胫部和跗跖部一般为角质鳞片所覆盖,且不具羽毛,以利其涉水行走;趾间有时具蹼,但与游禽不同,涉禽的足蹼仅存在于趾间基部,称为半蹼或微蹼,主要作用是增加与地面的接触面积,利于在湿地上行走;有些种类(如秧鸡、水雉)的脚趾细长,能在莲叶或浮萍上疾走。
鹳形目和鹤形目鸟类为大型涉禽,两者的外形及生活习性相近,以鱼、蛙等大型水生生物为食,嘴粗壮而锐尖,有如鱼叉,捕食方式是以静伺或潜行啄捕为主;或嘴端扁平有如汤匙 (如琵鹭) , 觅食时迎着水流将嘴尖插入水内,左右晃动头部来搜索猎物。鹳形目鸟类的后趾发育,能栖树握枝,而鹤类后趾退化,从不树栖,不能握枝。
鸻形目鸟类为中小型涉禽,是一群被称之为滨鸟的湿地水鸟, 包括鸻、 鹬、滨鹬、杓鹬、塍鹬、反嘴鹬、长脚鹬、蛎鹬、沙锥、翻石鹬和麦鸡等主要类型。它们生活在潮汐港湾的泥地和湿地上,通常具长腿、尖喙和长而尖的翅。它们的嘴较细弱,而且随取食方式不同有多种形态,主要在海岸的潮间带或河、湖的岸边啄食螺类等小型水生生物,边觅食边行走,十分迅捷。其觅食方式分触觉性觅食和视觉性觅食。典型的视觉性觅食方式为“啄—奔—啄” ,通过快速奔跑再一个急停,取食滩涂表面来不及躲入泥沙深处或浅水中的底栖生物,并有单啄、翻动啄食和多次啄食等几种形式。这种觅食方式受到个体间的觅食干扰很大,因此它们在觅食时尽可能地减少个体间干扰,而行单独或低密度分散觅食(以鸻类为主) 。触觉性取食是将长喙插入泥中,靠喙对食物的感觉来探搜觅食,有单次探搜和多次探搜等形式。觅食时,它们的运动距离较短,长喙深入泥中探搜较长时间才抬头转移。因此这类水鸟觅食时个体间相互干扰少,可成群觅食(以鹬类为主) 。
涉禽的尾大多较短,大型种类的翅长而宽,可做短距离的翱翔;小型种类(如鸻鹬类)翅短而尖,飞行迅速而灵活,体羽多数以灰、褐为主,与沙滩泥沼的颜色十分接近,是有效的保护色。
涉禽一般在树上、 草丛中或岩缝、 屋顶上(如鹳类)或浅水地带草丛中(如鹤类)营巢,以树枝及草茎编巢;有些种类的巢十分简陋,一般是在水边地面挖一浅凹,不加巢材铺垫(如鸻鹬类) ,但它们卵的颜色与环境相似,很难被发现。不少涉禽可以在距离湿地较远的树林中集群营巢,甚至可以混群集群营巢,形成极为壮观的湿地水鸟营巢地(如鹭类) 。鹳形目鸟类的雏鸟为晚成鸟;鹤形目和鸻形目的雏鸟属于早成鸟。
涉禽大多为迁徙性鸟类,它们通常从一个半球飞到另外一个半球。与游禽一样,涉禽的大多数在北半球繁殖,秋季南迁到比较温暖的湿地区越冬,第二年春季返回北方繁殖地。当然也有一些种不做长距离迁徙,只进行中等距离的迁徙,以便找到其适合的栖息地,但是它们不采用季节性迁徙的方式。